Влияние возраста женщины на процесс оплодотворения ооцитов и развитие эмбрионов в культуре
|
Одним из значительных достижений уходящего столетия без сомнения можно считать изменение социального положения женщин во многих странах мира. Получение образования, карьера, достижение определенного общественного статуса потребовали от них изменения отношения к такому важнейшему предназначению, как продолжение рода. Откладывание рождения ребенка на более поздний период жизни привело к тому, что к моменту, когда женщина решается на этот шаг, ее возможности в репродукции весьма ограничены. Бесплодие является другой причиной, по которой женщины старшего возраста продолжают поиск путей к рождению ребенка. Решение этих проблем предлагается врачам и биологам.
ЭКО открыло новые возможности в области репродукции. С помощью этого метода стало реальным не только преодолеть бесплодие, но и подойти к изучению ряда биологических феноменов с новых позиций. Не все из известных методов оценки гамет и эмбрионов могут быть использованы в клинической программе ЭКО в полной мере. Так, детальное исследование морфологических характеристик яйцеклетки невозможно из-за опасности ее повреждения. По этой причине о качестве яйцеклетки судят по ее функциональным характеристикам, т.е. способности к нормальному оплодотворению и дальнейшему развитию. При этом исходят из того, что первые деления дробления регулируются факторами яйцеклетки, а влияние мужского генома начинает проявляться на стадии 4—8 бластомеров [1]. В этой связи исследование особенностей оплодотворения ооцитов и последующего развития эмбрионов можно рассматривать как способ оценки ооцитов. Данные литературы о влиянии возраста женщины на процессы оплодотворения яйцеклеток и развитие эмбрионов человека в культуре достаточно противоречивы. M.Hull и соавт. [2] при анализе результатов 561 процедуры ЭКО у женщин в возрасте от 25 до 44 лет с первой попыткой лечения, имевших овуляторный цикл и нормальные показатели спермограммы мужа, не выявили зависимости частоты оплодотворения ооцитов от возраста пациенток. По мнению американских исследователей возраст не влияет на процессы фрагментации и число бластомеров у эмбрионов [3]. Этими авторами отмечено, что единственным морфологическим признаком эмбрионов, который при этом изменяется, является толщина блестящей оболочки. В то же время A.Stolwijk и соавт. [4] показали, что с возрастом уменьшается число перенесенных эмбрионов с нормальной морфологией. Задачей настоящей работы явилось изучение влияния возраста женщины на процессы оплодотворения, характер формирования зигот, темп дробления эмбрионов, степень фрагментации бластомеров. Материал и методы Проанализированы результаты оплодотворения ооцитов в культуре у 774 женщин, проходивших лечение методом ЭКО в нашем центре. Показанием к ЭКО у всех больных явилось трубно-перитонеальное бесплодие вследствие непроходимости или отсутствия маточных труб. Возраст больных варьировал от 20 до 46 лет и составлял в среднем 30,8±0,1 года. Показатели спермограммы у их мужей были нормальными. Стимуляцию суперовуляции проводили с использованием кломифенцитрата (клостилбегит) и человеческого менопаузального гонадотропина (хумегон, пергонал). Для индукции овуляции вводили хорионический гонадотропин при достижении лидирующими фолликулами диаметра 17—18 мм. В среднем было пунктировано 7,74±0,14 фолликула и получено 5,58±0,11 ооцита. Ооциты отмывали и культивировали в среде Ham-F10 с альбумином человека в течение 6 ч. Для инсеминации вводили 100 000 сперматозоидов в 1 мл среды. Через 18 ч определяли число пронуклеусов в зиготах, при этом полиплоиды удаляли, а нормальные зиготы переносили в свежую среду. Через 24 ч с момента контроля оплодотворения (первая оценка) эмбрионы анализировали под микроскопом и определяли число бластомеров. Выделяли три основные типа эмбрионов — без фрагментации бластомеров, а также с легкой и тяжелой степенью фрагментации цитоплазмы. Эмбрионы первой группы характеризовались индексом фрагментации 3, второй — 2 и третьей — 1. Вторая оценка развития эмбрионов происходила перед переносом их в полость матки через 48 ч с момента контроля оплодотворения ооцитов. Для переноса отбирали 4 эмбриона с наибольшим числом бластомеров и наименьшей степенью фрагментации цитоплазмы. Всего проанализировано 2760 эмбрионов. Частоту оплодотворения вычисляли как отношение числа зигот к общему числу полученных ооцитов. Частоту дробления зигот определяли как отношение числа эмбрионов к общему числу полученных зигот. Частоту встречаемости полиплоидных зигот вычисляли по отношению числа полиплоидных зигот к общему числу ооцитов. Для анализа полученных данных использовали методы вариационного, дисперсионного, корреляционного и регрессионного (параметрического линейного, а также множественного) статистического анализа. Результаты В среднем при оплодотворении в культуре было получено 3,6±0,08 эмбриона, после отбора прогрессивно развивающихся было перенесено в полость матки 2,8±0,04 эмбриона. Частота оплодотворения в среднем составила 72,1±0,8%, частота дробления зигот — 94,3±0,5%, частота полиплоидных зигот — 5,5±0,4%. При использовании корреляционного анализа в группе пациенток от 20 до 39 лет не было выявлено зависимости частоты оплодотворения ооцитов от возраста женщины. Частота оплодотворения ооцитов в культуре была снижена только у женщин старше 39 лет. В данной возрастной группе этот показатель составил 61,5±8,4%, что достоверно ниже, чем у пациенток в возрасте 20—24 года (82,4±3,4%; р меньше 0,05 ), а также 35—39 лет (80,8±2,1%; р меньше 0,05). Частота дробления и частота образования полиплоидных зигот не зависели от возраста женщины (табл. 1). Таблица 1. Частота развития зигот и полиплоидии (в %) у женщин разных возрастных групп
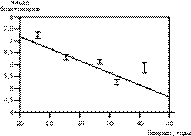
Замедленный темп дробления, связанный с возрастом пациенток, отмечен нами при оценке эмбрионов через 24 ч (r = –0,08; р меньше 0,0001) и 48 ч (r = –0,11; р меньше 0,0001) после контроля оплодотворения. Развитие лучших эмбрионов, выбранных впоследствии для переноса в полость матки, проходило с той же закономерностью (табл. 2). При первой оценке коэффициент регрессии равнялся –0,096 (р меньше 0,0001), при второй составил –0,14 (р меньше 0,0001). На рис. 1 приведена математическая модель зависимости числа бластомеров (при оценке всех полученных эмбрионов через 48 ч после контроля оплодотворения) от возраста женщины. Эта зависимость носит линейный характер и свидетельствует о постепенном закономерном снижении числа бластомеров с возрастом.
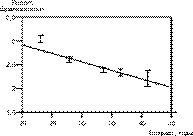
Достоверное повышение степени фрагментации с возрастом наблюдалось при оценке эмбрионов через 48 ч как среди всех полученных эмбрионов (r = –0,096; р меньше 0,0001), так и среди эмбрионов, перенесенных в полость матки (r = –0,014; р меньше 0,00001) (см. табл. 2). Повышение степени фрагментации (снижение индекса фрагментации) также носит линейный характер (рис. 2).
Обсуждение На большой выборке при использовании высокоинформативных методов статистического анализа нами установлено, что параметры, характеризующие способность ооцитов к оплодотворению, не имеют существенных различий у женщин в возрасте от 20 до 39 лет. Снижение частоты оплодотворения яйцеклеток носит закономерный характер в группе женщин старше 39 лет. В то же время достаточно рано возраст женщин начинает сказываться на темпе дробления эмбрионов и на степени выраженности процесса фрагментации. По всей видимости, способность к оплодотворению у яйцеклетки остается ненарушенной дольше, чем сохранность механизмов, обеспечивающих нормальное развитие эмбриона. Одной из возможных причин, приводящих к нарушению развития эмбрионов у женщин старшей возрастной группы, может являться повышение уровня хромосомных аберраций в ооцитах. Так, нами была выявлена тенденция к увеличению числа гетероплоидных женских половых клеток с возрастом [5]. Эти данные совпадают с результатами работы, выполненной D.Battaglia и соавт. [6] на модели созревания ооцитов в культуре, в которой показано, что частота хромосомных аберраций в этих клетках у женщин в возрасте 40—45 лет составляет до 79%, а у молодых женщин (20—25 лет) — 17%. Аналогичные результаты были получены K.Volarcik и соавт. [7]. Частота хромосомных аберрации в ооцитах у женщин моложе 35 лет составляла 21%, а в возрасте 35 лет и старше — 71%. Такой высокий уровень хромосомных аберраций в ооцитах может приводить к нарушению эмбриогенеза. Известно, что до 60% эмбрионов, развитие которых блокируется на доимплантационной стадии (с сильной степенью фрагментации бластомеров) являются генетически неполноценными: анеуплоидными или мозаичными [8, 9]. Выводы Частота оплодотворения ооцитов не изменяется у женщин в возрасте от 20 до 39 лет и постепенно снижается у женщин старше 40 лет. Частота формирования полиплоидных зигот и развития диплоидных не зависит от возраста. В то же время наблюдается постепенное снижение скорости дробления эмбрионов и повышение частоты их фрагментации. О.А.Воробьева, В.С.Корсак, Е.Л.Северова, А.А.Кирсанов, В.В.Козлов НИИ акушерства и гинекологии им. Д.О.Отта РАМН, Международный центр репродуктивной медицины, Санкт-Петербург Литература 1. Braude P., Bolton V., Moore S. Human gene expression first occurs between the four and eight-cell stages of preimplantation development. Nature 1988; 332: 459–461. 2. Hull M.G., Fleming C.F., Hughes A.O., McDermott A. The age-related decline in female fecundity: a quantitative controlled study of implanting capacity and survival of individual embryos after in vitro fertilization. Fertil Steril 1996; 65: 783–790. 3. Alikani M., Wiemer K. Embryo number for transfer should not be strictly regulated. Fertil Steril 1997; 68: 782–783. 4. Stolwijk A., Zielhuis G.A., Sauer M.V., Hamilton C.J., Paulson R.J. The impact of the womans age on the success of standard and donor in vitro fertilization. Fertil Steril 1997; 67: 702–710. 5. Воробьева О.А., Кузнецова Т.В., Богомолова М.В., Баранов В.С. Анализ частоты хромосомных аберраций в ооцитах человека. Пробл репрод 1997; 2: 21–26. 6. Battaglia D.E., Goodwin P., Klein N.A. Influence of maternal age on meiotic spindle assembly in oocytes from naturally cycling women. Hum Reprod 1996; 11: 2217–2222. 7. Volarcik К., Sheean L., Goldfarb J., Woods L., Abdul-Karim F., Hunt P. The meiotic competence of in vitro matured human oocytes is influenced by donor age: evidence that folliculogenesis is compromised in the reproductively aged ovary. Hum Reprod 1998: 13: 154–160. 8. Janny I., Menezo Y. Maternal age effect on early human embryonic development and blastocyst formation. Mol Reprod Dev 1996; 45: 31–37. 9. Munne S., Rosenwaks Z., Grifo J., Cohen J. Diagnosis of major chromosome aneuploidies in human preimplantation embryos. Hum Reprod 1993; 8: 2185–2191. 10. Munne S., Grifo J., Cohen J., Weier H.-U.G. Chromosome abnormalities in human arrested preimplantation embryos: a multiple-probe FISH study. Am J Hum Genet 1994; 55: 150–159. |
||||||||||||||||||||||||||